Metabolism
on-line - the virtual tutorial room
copyright © 2008 - 2015 David A Bender
Is fructose fattening?
One hypothesis to explain the increase in overweight and obesity over the last two decades is that as well as increased food consumption and reduced physical activity, increased consumption of fructose may be a factor. Fructose is 1.7-times sweeter than sucrose, and high fructose syrups, made by hydrolysis of starch followed by isomerisation of much of the glucose to fructose, provide a cheap and convenient alternative to sucrose in the manufacture of sweetened beverages and other foods. There is good experimental evidence in experimental animals, and some preliminary evidence in human beings, that consumption of a relatively large amount of fructose leads to increased fatty acid and triacylglycerol synthesis in the liver, and increased secretion of very low density lipoprotein. Consumption of an equivalent amount of glucose is not associated with the same increase in lipogenesis.
Which step provides the main regulation of glycolysis?
The main control over the rate of glycolysis is by regulation of the activity of phosphofructokinase.

What compounds would you expect to inhibit phosphofructokinase?
Phosphofructokinase is inhibited by three compounds:
ATP (although ATP is a substrate, there is a separate inhibitory ATP-binding site on the enzyme)
phosphoenolpyruvate
citrate
The graphs below show the effect of ATP on the substrate dependence of phosphofructokinase, and the inhibition of phosphofructokinase by ATP in the presence and absence of 5'AMP.

What is the metabolic significance of the inhibition of phosphofructokinase by physiological concentrations ATP?
ATP can be considered to be the most important end-product of glycolysis and the citric acid cycle. Inhibition of phosphofructokinase by ATP thus represents feed-back inhibition of the main controlling enzyme of the pathway. When there is enough ATP in the cell there is no need to produce any more (indeed, it would not be possible, since the rate of oxidation of metabolic fuels is controlled by the availability of ADP to be phosphorylated, see the exercise on Overheating after overdosing on E - and slimming by taking dinitrophenol)
What is the effect of 5'AMP on phosphofructokinase, and what is its significance?
At normal intracellular concentrations of ATP and fructose 6-phosphate the enzyme is almost 90% inhibited. However, in the presence of a relatively small amount of 5'AMP this inhibition is more or less completely reversed, so that the rate of glycolysis increases 10-fold, so permitting more phosphorylation of ADP to ATP.
![]()
5'AMP is formed by the reaction of adenylate kinase, shown above. There is normally very little ADP present in cells. However, in response to muscle contraction and other uses of ATP, ADP is formed, but not enough to produce a significant change in the ration of ATP : ADP, which is about 500 : 1. The small amount of 5'AMP that is formed by the adenylate kinase reaction acts as a potent signal of the energy state of the cell, increasing the rate of glycolysis (and other energy-yielding reactions) very considerably.
What is the metabolic significance of inhibition of phosphofructokinase by phosphoenolpyruvate?
Phosphoenolpyruvate is an intermediate in glycolysis, but is rapidly converted to pyruvate by pyruvate kinase. However, when gluconeogenesis is stimulated by the hormone glucagon (in response to a low blood concentration of glucose), pyruvate kinase is inactivated by phosphorylation of the enzyme protein.
More importantly, phosphoenolpyruvate is synthesised from oxaloacetate as a key reaction in gluconeogenesis. It is obviously essential that if ATP is being expended to synthesise glucose from non-carbohydrate precursors (e.g. in the fasting state), then the reactions of glycolysis must be inhibited - hence the inhibition of phosphofructokinase by phosphoenolpyruvate.

What is the metabolic significance of inhibition of phosphofructokinase by citrate?
Citrate can also be considered to be an end-product of glycolysis; accumulation of citrate in the cytosol implies that sufficient acetyl CoA is being formed to maintain citric acid cycle activity and ATP production.
What we need to consider is how the efflux of citrate from the mitochondrion is controlled, so as not to deplete citric acid cycle intermediates and so impair ATP production.
Equally, we have to explain how acetyl CoA is available in the cytosol for fatty acid synthesis - the only pathways that form acetyl CoA are in the mitochondrion, and acetyl CoA does not cross the mitochondrial membrane.
The only source of acetyl CoA is inside the mitochondrion, but acetyl CoA is required in the cytosol for fatty acid synthesis.
Citrate is formed in the mitochondrion by reaction of acetyl CoA with oxaloacetate. When acetyl CoA is required for fatty acid synthesis, citrate is transported out into the cytosol, where it is cleaved by citrate lyase, forming acetyl CoA that can be used for fatty acid synthesis, and oxaloacetate.
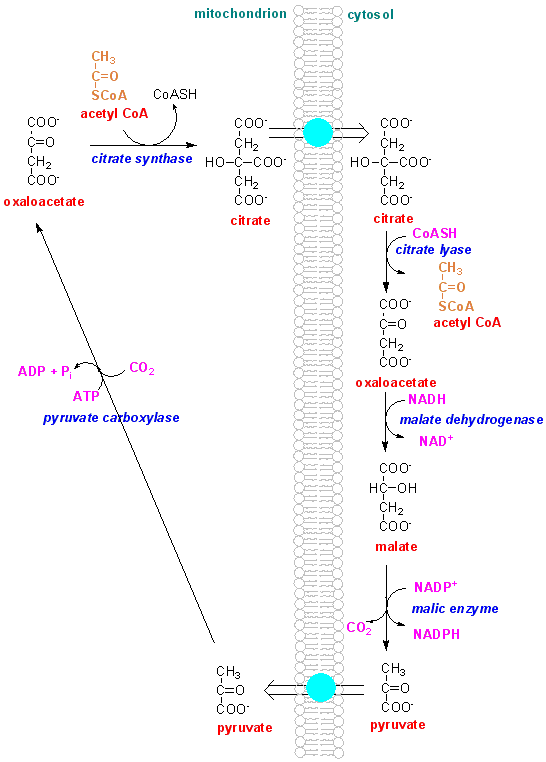
What will be the main source of acetyl CoA in the mitochondria under conditions when fatty acids are to be synthesised?
Fatty acids are synthesised in the fed state, when the main source of acetyl CoA will be pyruvate, the end-product of glycolysis. Several amino acids that are surplus to requirements for protein synthesis give rise to pyruvate; others give rise of acetyl CoA.
There will be no acetyl CoA arising from either fatty acids or ketone bodies. In the fed state ketone bodies are not synthesised to any significant extent, and fatty acid transport into the mitochondrion is strongly inhibited by malonyl CoA, which is formed by carboxylation of acetyl CoA as the main substrate for fatty acid synthesis (see the exercise on Do we need to synthesise fatty acids?).
Oxaloacetate cannot re-enter the mitochondrion, but undergoes reduction to malate and oxidative decarboxylation to pyruvate (see also the exercise on How is NADH from glycolysis normally re-oxidised? and compare the pathway here with the malate-aspartate shuttle for transferring reducing equivalents form NADH in the cytosol to NADH in the mitochondrion).
Pyruvate enters the mitochondrion, and can be carboxylated to oxaloacetate.
What is the metabolic advantage of the pathway by which oxaloacetate is reduced to malate by malate dehydrogenase, then oxidised (and decarboxylated) to pyruvate by the malic enzyme?
There is a requirement for 2 x NADPH for each mol of acetyl CoA incorporated into the growing fatty acid (see the exercise on Do we need to synthesise fatty acids?).There is an ample supply of NADH in the cytosol when glycolysis is occurring, and this reaction permits production of NADPH at the expense of NADH.
The malic enzyme only provides one mol of NADPH for each acetyl CoA transported out of the mitochondrion, but two are required.
What is the source of other half of the NADPH that is required for fatty acid synthesis?
The rest of the NADPH that is required for fatty acid synthesis comes from the pentose phosphate pathway (see the exercise on An adverse response to antimalarial medication - and a fatal reaction to fava beans).
In a series of experiments, rat mammary gland tissue was incubated with [14C-1] glucose or [14C-6] glucose. Tissue was taken from lactating animals (which synthesise a large amount of fatty acid in the mammary gland to be secreted into the milk) and non-lactating animals (which synthesise little or no fatty acid in the mammary gland. The results were as follows (figures show radioactivity in carbon dioxide, expressed as dpm /mg protein in a 20 minute incubation, ± sd for 5 x replicate incubations):
[14C-1]glucose |
[14C-6]glucose |
|
| lactating mammary gland | 3406 ± 95 |
1208 ± 78 |
| non-lactating mammary gland | 2103 ± 87 |
2008 ± 91 |
What conclusions can you draw from these results?
When glucose 6-phosphate is metabolised in thepentose phosphate pathway, carbon-1 of glucose is lost as carbon dioxide. When glucose 6-undergoes complete oxidation in glycolysis and the citric acid cycle the amount of carbon dioxide formed from carbon-1 and carbon-6 of glucose will be equal, because of the rapid isomerisation of glyceraldehyde 3-phosphate and dihydroxyacetone phosphate. Therefore, if there is significantly more radioactive carbon dioxide from [14C-1]glucose than from [14C-6]glucose, this suggests that there is increased activity of the pentose phosphate pathway.
What is the metabolic disadvantage of the pathway by which oxaloacetate is reduced to malate by malate dehydrogenase, then oxidised (and decarboxylated) to pyruvate by the malic enzyme?
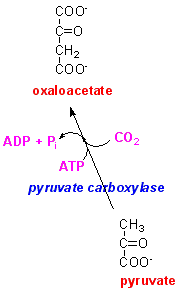 There
is a cost of 1 x ATP for each mol of pyruvate that has to be carboxylated to
oxaloacetate to maintain an adequate concentration of oxaloacetate in the mitochondrion
for citric acid cycle activity.
There
is a cost of 1 x ATP for each mol of pyruvate that has to be carboxylated to
oxaloacetate to maintain an adequate concentration of oxaloacetate in the mitochondrion
for citric acid cycle activity.
Isolated hepatocytes were incubated for 40 min in a phosphate/bicarbonate buffer
system, with
[14C-U]palmitate (i.e., the C16 fatty acid, palmitate, labelled
with 14C in all 16 carbon atoms), at a specific radioactivity of 1000 dpm /mmol,
with and without the addition of 60 mmol/L malonate (which blocks the citric
acid cycle by inhibiting succinate dehydrogenase) and / or malate.
The reaction was stopped by the addition of perchloric acid to denature proteins and drive off carbon dioxide, which was trapped in an organic base to permit determination of radioactivity. The denatured incubation mixture was then extracted with a chloroform:methanol mixture to separate unmetabolised fatty acid (palmitate, in the organic phase) from water-soluble metabolites (in the aqueous layer). The radioactivity in both phases was determined.
One set of incubations was set up to act as a control, in which perchloric acid was added at the beginning of the experiment to denature proteins and drive off carbon dioxide.
The results were as follows (figures show dpm /min /gram cells, ± sd for 5 x replicate incubations)
carbon dioxide |
organic phase |
aqueous phase |
|
| unincubated control | 0 |
9989 ± 120 |
0 |
| incubated, no addition | 2300 ± 120 |
7526 ± 98 |
215 ± 21 |
| incubated + malonate | 0 |
9985 ± 105 |
205 ± 19 |
| incubated + malate | 4800 ± 230 |
4975 ± 110 |
226 ± 18 |
| incubated + malate + malonate | 0 |
5012 ±120 |
5036 ± 110 |
What conclusions can you draw from these results?
The incubation with added malate shows the catalytic effect of adding additional"carrier" for the acetyl CoA, so that it is oxidised to carbon dioxide faster - there is more radioactive carbon dioxide formed in the incubation with added oxaloacetate than with no additions.
For each two carbon atoms that enter the citric acid cycle as acetyl CoA, forming citrate, two carbon atoms are lost as carbon dioxide in the reactions that lead to the reformation of the 4-carbon compound oxaloacetate. However, when malonate is added there is no radioactivity in carbon dioxide, even when malate is also added as a source of oxaloacetate that can enter the mitochondrion.
When both malonate and malate are added there is a considerable accumulation of a water soluble metabolite.

What do you think this water-soluble metabolite is?
This will be mainly succinate, the substrate of the enzyme that has been inhibited by the added malonate. Other intermediates (citrate, isocitrate, ketoglutarate and succinyl CoA) will also be labelled - when no malonate is added, fumarate, malate and oxaloacetate will also be labelled.
Why do you think that no radioactive carbon dioxide is produced in the presence of malonate, even when malate is also added?
Obviously, the two carbon atoms that were added from acetyl CoA to form citrate are not the same two atoms as are lost as carbon dioxide in the first turn of the cycle. In the absence of malonate, all of the intermediates of the cycle become labelled, and in the second and subsequent turns of the cycle there is radioactivity in the carbon dioxide that is lost.
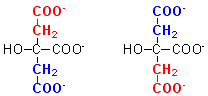 This
raises a further problem. Although the results above clearly show that the two
carbon atoms added from acetyl CoA, which become carbons 1 and 2 of citrate,
are not lost as carbon dioxide in the first turn of the cycle, citrate is a
symmetrical molecule, and carbons 1 and 2 are equivalent to carbons 5 and 6.
This
raises a further problem. Although the results above clearly show that the two
carbon atoms added from acetyl CoA, which become carbons 1 and 2 of citrate,
are not lost as carbon dioxide in the first turn of the cycle, citrate is a
symmetrical molecule, and carbons 1 and 2 are equivalent to carbons 5 and 6.
How do you think a symmetrical molecule like citrate can behave asymmetrically?
Citrate must remain enzyme bound, and not go into free solution. This means that it must be channelled directly from the catalytic site of citrate synthase to that of aconitase. Isocitrate, the product of the aconitase reaction, is asymmetric.
It is only when the active site of aconitase is occupied that citrate can leave citrate synthase to go into free solution. This means that citrate will only leave the mitochondrion when the citric acid cycle is running as fast as is required for ATP synthesis. If there is a need for citrate to undergo oxidation for ATP production, the active site of aconitase will not be occupied, and citrate can be passed directly from citrate synthase to aconitase.
![]() In
the next set of experiments, hepatocytes were incubated with [14C-3]pyruvate,
together with malonate to inhibit succinate dehydrogenase, malate as a source
of oxaloacetate, and phenylacetic acid to inhibit pyruvate carboxylase.
In
the next set of experiments, hepatocytes were incubated with [14C-3]pyruvate,
together with malonate to inhibit succinate dehydrogenase, malate as a source
of oxaloacetate, and phenylacetic acid to inhibit pyruvate carboxylase.
The specific activity of the [14C-3]pyruvate was 1 µCi /µmol.
What would you expect to be the specific activity of the palmitate formed?
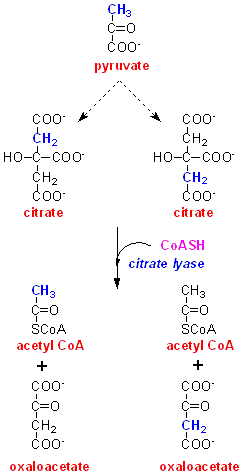 At
first glance you might expect the specific activity of palmitate to be 8 µCi
/µmol, since 8 x acetyl CoA, derived from pyruvate by way of citrate synthesis
and cleavage, will be required for palmitate synthesis. However, citrate is
a symmetrical molecule, and when it leaves the mitochondrion carbon-2, which
has arisen from carbon-3 of pyruvate, is equivalent to carbon-5.
At
first glance you might expect the specific activity of palmitate to be 8 µCi
/µmol, since 8 x acetyl CoA, derived from pyruvate by way of citrate synthesis
and cleavage, will be required for palmitate synthesis. However, citrate is
a symmetrical molecule, and when it leaves the mitochondrion carbon-2, which
has arisen from carbon-3 of pyruvate, is equivalent to carbon-5.
This means that half the acetyl CoA formed by the action of citrate lyase will contain labelled carbon from carbon-3 of pyruvate, and half will contain unlabelled carbon from the malate that was added.
The specific activity of the palmitate formed will therefore be 4 µCi /µmol, half of what you would expect if citrate were not symmetrical.
What would you expect if you incubated the hepatocytes with [14C-2]malate at a specific activity of 1 µCi /µmol, and unlabelled pyruvate, together with malonate to inhibit succinate dehydrogenase and phenylacetic acid to inhibit pyruvate carboxylase?
Carbon-2 of malate becomes carbon-5 of citrate as it is formed on the active site of citrate synthase. Again, once citrate leaves the enzyme carbon-5 is equivalent to carbon-2, and so again you would expect the specific activity of palmitate to be 4 µCi /µmol.
Most fructose is metabolised by way of formation of fructose 1-phosphate, catalysed by fructokinase.

Looking at the pathway above, can you explain why a high intake of fructose will lead to increased synthesis of fatty acids?
As we have already seen, the main regulatory step of glycolysis is catalysed phosphofructokinase. However, as shown above, fructose metabolism via fructokinase leads to the formation of dihydroxyacetone phosphate and glyceraldehyde 3-phosphate directly, by-passing phosphofructokinase.
This means that glycolysis from fructose will not be controlled by ATP, phosphoenolpyruvate or citrate, resulting in the formation of pyruvate and then acetyl CoA in excess of the needs for ATP-yielding metabolism. The only metabolic fate available to this "surplus" acetyl CoA is condensation to form citrate, which is no needed for oxidative metabolism, and so leaves the mitochondrion and provides acetyl CoA for fatty acid synthesis.
Summarise the key points from this exercise